
La naissance de la biologie moléculaire |
Le XXe siècle coïncide avec la naissance
génétique : débutant avec la redécouverte
des travaux de Mendel précisément en 1900, se
poursuivant par l'élaboration de la théorie
chromosomique de l'hérédité au début du
siècle, la découverte de l'ADN comme support biochimique
de l'information génétique, l'élucidation de sa
structure, et l'explosion de la biologie moléculaire à
partir des années 70. Et l'aube du XXIe siècle verra
vraisemblablement le patrimoine génétique de l'homme
intégralement décrypté.
La génétique moderne remonte aux travaux de Mendel, qui le premier établit les lois de l'hérédité. Il publie ses résultats en 1866, mais ils passent alors à peu près inaperçus. Leur redécouverte n'aura lieu qu'en 1900.
Ce sont les travaux de Morgan, sur la drosophile, qui conduisent au développement de la théorie chromosomique de l'hérédité. Les gènes sont alors localisés sur les chromosomes, et avec Sturtevant, ils pourront même y être ordonnés, constituant les premières cartes génétiques. C'est encore dans le laboratoire de Morgan que sont développées les procédures de mutagenèse expérimentales par Muller.
Si la présence des gènes sur les chromosomes est alors établie, rien n'est connu de la nature biochimique des gènes ou de leur mode d'action. La première relation entre un gène et un enzyme est établie en 1902 par Garrod, à partir d'une observation portant sur une maladie génétique humaine. Beadle et Tatum approfondissent cette relation sur un système accessible à l'expérimentation, le champignon Neurospora crassa. L'ensemble de ces travaux aboutissent finalement à la conclusion que les gènes contrôlent la synthèse des enzymes, et que chaque protéine est codée par un gène différent.
Le premier phénomène qui allait permettre de progresser dans l'identification du support de l'hérédité est celui de la transformation bactérienne, rapporté en 1928 par l'anglais Griffith . Ce phénomène représente alors un test d'activité biologique, grâce auquel il est possible de déterminer la nature du matériel génétique. Ce test ne sera pas mis à profit par Griffith lui même, mais par Avery qui l'utilise pour élucider la nature biochimique du matériel génétique : il s'agit de l'ADN. Cette découverte est toutefois accueillie avec beaucoup de scepticisme. Il faudra de nombreux autres travaux pour que cette réalité soit acceptée : en particulier ceux de Chargaff ou de Hershey. L'acceptation définitive ne viendra qu'avec l'élucidation de la structure de l'ADN par Watson et Crick.
L'influence des physiciens va marquer la génétique
moléculaire. Certains, comme Schrödinger, n'ont qu'un rôle
d'observateur. D'autres y consacrent leur carrière, car il
perçoivent cette science comme la nouvelle frontière de
la connaissance scientifique. Ainsi Pauling ou
Delbrück jouent un rôle
déterminant dans le développement de cette
science. Delbrück sera en particulier le fondateur du Groupe du
phage, avec Luria et
Finalement, c'est avec l'élucidation de la structure de l'ADN que la biologie moléculaire connaît son apothéose. Cette réussite est le fait de Watson et Crick, mais aussi de chercheurs tels que Franklin ou Wilkins.
 Gregor
Mendel (1822 - 1884)
Gregor
Mendel (1822 - 1884)La génétique classique débute avec les travaux d'un moine tchèque : Gregor Mendel, qui travaille de façon isolée dans un monastère morave. Il avait reçu au cours de sa formation les enseignements de Franz Unger (1800 - 1870) et Christian Doppler (1803-1853). Le premier est son professeur de botanique, qui avait déjà proposé des idées évolutionnistes, en avance sur son époque. Le second est son professeur de physique (c'est le découvreur de l'effet Doppler), dont l'influence s'est révélée déterminante pour l'analyse statistique que Mendel mène sur les résultats de ses croisements.
Grâce à de rigoureuses observations menées sur les pois (Pisum sativum), il établit dès 1866 les premières lois de l'hérédité. L'espèce choisie permet de mener à volonté soit des auto-fécondations, soit des fécondations croisées, et sa fécondité importante autorise des analyses sur un nombre important de plantes à chaque génération. Sur ce modèle expérimental, Mendel étudie la transmission au cours des générations d'un certain nombre de caractères simples à observer. Ce sont toujours des caractères à versions alternatives tranchées : fruits lisses ou rugueux, verts ou jaunes, graines rondes ou irrégulières, jaunes ou vertes, tige haute ou petite.
Grâce à l'autofécondation pratiquée sur de nombreuses générations, Mendel établit d'abord des lignées pures, dont la descendance présente toujours les mêmes caractères. A partir du croisement de ces lignées, l'analyse de la descendance obtenue allaient amener Mendel à formuler les trois premières lois de l'hérédité:
1) dans la première génération (génération F1) d'un croisement impliquant deux lignées pures différant par un unique caractère (monohybridisme), tous les individus présentent un même phénotype. Le caractère qui se manifeste à l'exclusion de l'autre dans la génération F1 est qualifié de dominant, et le caractère qui en est exclus est qualifié de récessif
2) dans la descendance d'un croisement impliquant deux individus F1 (génération F2), les deux caractères parentaux réapparaissent suivant une proportion prédictible de 3 à 1
3) si l'on croise des lignées pures différant pour plusieurs caractères (polyhybridisme), chacun de ces caractères se comporte de façon indépendante vis à vis de l'autre. Ainsi, pour un croisement impliquant deux caractères, les proportions observées sont 9:3:3:1.
Ces lois ont été amplement vérifiées par la suite. Elles ne sont toutefois pas d'utilisation universelle. D'une part, Mendel n'explique pas l'existence, pour certains caractères, d'individus F1 présentant un phénotype intermédiaire entre celui des parents (du à une codominance ou un polygénisme). D'autre part, elles ne s'appliquent pas aux gènes liés sur un même chromosome (c'est une chance extraordinaire que Mendel se soit attaché à l'étude de gènes portés par des chromosomes différents ou suffisamment éloignés l'un de l'autre). Au début du siècle suivant, la mise en évidence de caractères liés allait conduire Morgan à proposer la théorie chromosomique de l'hérédité .
Les lois de Mendel impliquent l'existence d'éléments autonomes et reproductibles, qui contrôlent de façon discrète les caractères héréditaires de génération en génération. Chaque caractère est représenté dans l'œuf fécondé par deux - et seulement deux - éléments, provenant l'un du père, l'autre de la mère. Les autres théories de l'hérédité proposées par des contemporains de Mendel (en particulier Darwin, Weismann, de Vries ou Galton) postulaient toutes la présence simultanée de nombreux éléments déterminant un caractère donné dans chaque cellule.
Enfin, les travaux de Mendel réfutent la théorie de l'hérédité par mélange, théorie alors largement acceptée qui propose que les déterminants d'un caractère donné fusionnent après fécondation. En effet, Mendel n'observe pas de transition graduelle entre les caractères parentaux, et l'intégrité de chaque caractère est préservée lorsqu'ils réapparaissent en F2. Chez les hybrides F1, les éléments correspondant aux versions alternatives d'un caractère donné restent donc distincts, et se séparent à nouveau lors de la formation des cellules germinales. Ces particules, l'unité de l'hérédité, se verront attribuer par le biologiste danois Wilhem Johannsen (1857 - 1927) la dénomination de gènes en 1909.
Mendel publie ses résultats en 1866 dans un magistral article publié dans les Comptes rendus de la Société d'histoire naturelle de Brno. Ils passent alors à peu près inaperçus, n'étant cités qu'une douzaine de fois entre leur publication et leur redécouverte en 1900. Cette redécouverte est due à Hugo de Vries (Amsterdam), Carl Correns (Berlin), et Erich Tschermack (Vienne), qui retrouvent de façon indépendante des résultats similaires à ceux de Mendel.
Le fait que ces travaux soient restés si longtemps sans impact immédiat s'explique sans doute en partie par plusieurs lacunes qu'ils laissent transparaître pour les contemporains. Tout d'abord, les travaux de Mendel ne permettent pas d'expliquer l'atavisme, c'est à dire la réapparition d'un caractère qui existait non pas chez les parents ou grands-parents, mais chez un ancêtre qui peut être beaucoup plus éloigné. D'autre part, ces travaux ne concernent que les caractères à versions alternatives tranchées, et laissent complètement de côté les caractères à variations continues. La loi de Galton, bien qu'erronée, s'appliquait aux caractères à variations continues, et expliquait l'atavisme les lois : c'est cette théorie qui prévaut à l'époque.
Mayr, E. (1982) Histoire de la biologie (trad française : Fayard, 1989)
Fincham, J.R.S. (1990) Mendel - now down to the molecular level. Nature 343, 208-209.
Hugo de Vries est né à Haarlem, aux Pays-Bas. Entre 1878 et 1918, il est professeur de Botanique à Amsterdam. Il a à son actif plusieurs contributions importantes : la première est bien sûr la redécouverte des lois de Mendel en 1900 : alors qu'il croit mettre à jour certaines lois de l'hérédité, il s'aperçoit que Mendel l'a devancé de plus de 35 ans ! Sa seconde contribution concerne une théorie de l'hérédité impliquant des particules élémentaires qu'il baptise "pangènes". Cette théorie est plus proche de la réalité que toutes celles qui ont été proposées antérieurement.
Enfin, il développe une théorie de l'évolution par mutation : s'intéressant à la théorie de l'évolution proposée par Darwin, il cherche en particulier à déterminer si l'évolution est un phénomène graduel ou saltatoire. Pour cela, il met en culture un nombre considérable d'espèces de plantes herbacées, espérant ainsi observer chez l'une d'entre elles un changement brusque. C'est ce qu'il réussit finalement à observer chez Oenothera lamarckiana : parmi les descendants de cette espèce, un très petit nombre d'individus apparaissent présentant une variation discontinue. De Vries assimile alors ce phénomène à l'apparition d'une nouvelle espèce, et qualifie de mutation ce processus.
Il publie entre 1901 et 1903 La théorie de la mutation, livre où il prétend expliquer la naissance des espèces. D'après sa théorie, les espèces apparaissent en une seule génération, après qu'une variation de grande ampleur - une mutation - est apparue.
En réalité, l'observation rapportée chez Oenothera ne correspondait pas à une mutation génique au sens couramment admis aujourd'hui : ce type de variation est en fait imputable à des translocations chromosomiques, fréquentes dans cette espèce. Ce n'était pas non plus une nouvelle espèce qu'il observait, les individus décrits demeurant fertiles entre eux. De Vries eut cependant le mérite de mettre en évidence l'apparition de nouveautés génétiques.
L'influence de De Vries est considérable chez ses contemporains, c'est d'ailleurs en voulant valider cette théorie de la mutation chez la drosophile que Morgan sera amené à élaborer la théorie chromosomique de l'hérédité.
La théorie saltatoire de l'évolution présentée par de Vries se démarque notablement de la théorie gradualiste de Darwin, qui proposait que les nouvelles espèces apparaissent grâce à des modifications progressives : l'idée de Darwin était que la nature ne fait pas de saut (Natura non facit saltum). Les idées de de Vries vont cependant se développer dans le premier tiers du XXe siècle, avec le mouvement des évolutionnistes mutationistes (regroupant entre autres Bateson, Johannsen, ou Galton). Cette tendance était combattue par les évolutionnistes naturalistes, qui observaient les variations des espèces sur le terrain, et défendaient l'idée du gradualisme de l'évolution des espèces.
Le darwinisme revient cependant en force avec l'élaboration de la théorie synthétique de l'évolution entre 1936 et 1947. D'une part, les généticiens reconnaissent que les variations continues peuvent être dues à des facteurs mendéliens multiples, ce qui contribue à atténuer la contradiction entre la discontinuité des gènes et le caractère continu de la variation individuelle. D'autre part, les naturalistes adoptent progressivement la population comme unité de l'évolution. Les acteurs principaux de cette synthèse sont le naturaliste Ernst Mayr, le paléontologiste George Simpson, les généticiens Ronald Fisher, John Haldane, Alfred Sturtevant, Julian Huxley, et Theodosius Dobzhansky (la théorie synthétique de l'évolution sera cependant à nouveau contestée en 1968 par la théorie neutraliste de Motoo Kimura, et en 1972 par la théorie des équilibres ponctués de Niles Elderedge et Stephen Jay Gould).
Mayr, E. (1982) Histoire de la biologie (trad française : Fayard, 1989)
 Thomas Morgan (1866 - 1945)
Thomas Morgan (1866 - 1945)Morgan est né en 1866 à Lexington. Il obtient sa thèse en 1890 à l'université John Hopkins, sur une étude portant sur les Pycnogonides (arthropodes marins). Par la suite, il se tourne vers l'embryologie expérimentale : il s'intéresse en particulier à la régénération chez les vers et le développement de l'oursin. Il rejoint en 1904 l'université de Columbia, où se déroulera la phase la plus productive de sa carrière, portant sur la théorie chromosomique de l'hérédité. En 1928 il part au Cal Tech, où il restera jusqu'à sa mort en 1945.
Morgan avait été très intéressé par les travaux de de Vries sur les mutations, car ils proposaient un mécanisme permettant d'expliquer l'évolution et l'apparition de nouvelles espèces. Morgan se met alors lui aussi en quête de mutations de grande ampleur. Il choisit pour cela comme modèle d'étude la mouche du vinaigre (Drosophila melanogaster). Ce petit insecte se prête facilement à l'élevage et à l'observation, son cycle de reproduction est très court (9 jours à 25deg.C), il ne possède que 4 paires de chromosomes, et la descendance nombreuse à chaque génération (une femelle pond plusieurs centaines d'œufs).
La première mutation qu'il observe est une mouche mâle présentant des yeux blancs, au lieu d'yeux normalement rouges. Il remarque alors que ce caractère n'est jamais présenté que par les mâles, ce qui le conduit à proposer que le facteur déterminant ce caractère est porté par le chromosome sexuel. Ainsi, un facteur mendélien est pour la première fois expérimentalement assigné à un chromosome défini. Morgan définit ainsi l'hérédité liée au sexe.
Morgan est rejoint à partir de 1910 par Sturtevant. Ils vont identifier de nombreuses autres mutations, dont l'étude de la ségrégation va les conduire à établir les premières cartes génétiques,. Ces cartes sont complètement superposables aux chromosomes : toutes ces mutations se rangent en 4 groupes de liaison, correspondant à chacune des 4 paires de chromosomes. Ces travaux permettent donc de reconnaître les chromosomes comme support physique des gènes, et d'établir la théorie chromosomique de l'hérédité : les gènes sont organisés en série linéaire le long du chromosome. Ces découvertes furent résumées dans un livre publié en 1915 : Le mécanisme de l'hérédité mendélienne (Morgan, Sturtevant, Muller, Bridges). Pour ces travaux, Morgan reçut le prix Nobel de physiologie et de médecine en 1933.
Il convient de remarquer qu'une théorie similaire avait été précédemment avancée par les cytologistes Sutton (1903) et Boveri (1904), mais qu'elle n'avait pas alors reçut bon accueil. Morgan lui-même s'y était vigoureusement opposé entre 1903 et 1910, ce sont ses propres données expérimentales qui le conduisirent à se "convertir" à la théorie chromosomique de l'hérédité.
Le laboratoire de Morgan a été extraordinairement fécond, en raison d'un magnifique esprit de collaboration, resté légendaire. Plusieurs scientifiques de grande envergure y feront leurs premières contributions : outre Sturtevant et Bridges, ce laboratoire accueillît aussi Théodosius Dobzhansky, qui allait devenir un des maîtres d'œuvre de la théorie synthétique de l'évolution, et Hermann Muller, qui allait mettre au point en 1927 l'induction artificielle de mutations par les rayons X.
Morgan, T.H. (1910) Sex limited inheritance in Drosophila. Science 32, 120.
Morgan, T.H. (1911) The origin of nine wing mutations in Drosophila. Science 33, 496.
Morgan, T.H. (1911) The origin of five mutations in eye color in Drosophila and their mode of inheritance. Science 33, 534.
Morgan, T.H., Sturtevant, A.H., Muller, H.J. & Bridges, C. (1915) The mechanism of mendelian heredity. New-York.
 Alfred Sturtevant (1891 - 1970)
Alfred Sturtevant (1891 - 1970)Alfred Sturtevant est né en 1891 à Jacksonville. Il entre à l'université à 17 ans, et publie à 19 ans ses premiers travaux scientifiques. A partir de 1910, Sturtevant travaille avec Morgan à l'université de Columbia. Ils identifient chez la drosophile plusieurs mutations portant sur la couleur des yeux, la forme des ailes, la couleur du corps. Ils observent alors que tous ces caractères semblent liés au chromosome X. Les gènes qui les déterminent sont donc vraisemblablement portés par ce chromosome sexuel.
L'analyse de leur ségrégation montre que ces gènes sont en général transmis ensemble à la génération suivante. Cependant, dans de rares cas, ils semblent pouvoir être séparés. Sturtevant et Morgan rapprochent alors ces observations à celles du cytologiste belge Janssens, qui avait remarqué en 1909 que lors de la méiose, il se produit quelques fois des enjambements (crossing-over) entre chromosomes homologues dans la région qui les sépare. De tels crossing-over entraînent l'échange de portions de chromosomes entre les deux homologues impliqués. Toutes ces observations s'intègrent alors dans un schéma cohérent : les allèles portés par deux gènes portés sur le même chromosome sont séparés dans le cas où un crossing-over se produit dans l'intervalle qui les sépare.
Morgan et Sturtevant ont alors l'idée géniale de corréler la fréquence de tels événements à la distance qui sépare les gènes, et de proposer que la fréquence de recombinaison exprime une distance entre gènes. L'estimation des distances séparant les différents gènes allait conduire à l'établissement des premières cartes génétiques. Ces cartes sont établies en analysant la ségrégation des caractères correspondant au cours des générations, les distances reflétant le degré de liaison qui les lie. L'unité de distance adoptée est le centimorgan (cM) : un centimorgan correspond à une fréquence de recombinaison de 1% (un crossing-over pour cent méioses) entre deux marqueurs.
Sturtevant et Morgan publièrent en 1913 la première carte génétique jamais établie d'un chromosome, celle du chromosome X de la drosophile (Sturtevant est alors âgé de 22 ans). Elle montrait l'ordre et la succession des gènes y (corps de couleur jaune), w (yeux blancs), et m (ailes vestigiales).
The linear arrangement of six sex-linked factors in Drosophila, as shown by their mode of association. J. Exp. Zool. 14, 43-59.
Muller travaille tout d'abord avec Morgan à l'université de Columbia. Par la suite, à l'université du Texas, il met au point en 1927 l'induction artificielle de mutations par les rayons X : il montre que les rayons X augmentent d'un facteur 100 le nombre de mutations chez la drosophile.
Cette technique de production de mutation n'est pas spécifique, car une mutation ne peut être obtenue sur un gène choisi à l'avance. La possibilité d'augmenter la fréquence des mutations va cependant s'avérer d'un intérêt considérable : le nombre de mutations décrites va véritablement exploser, permettant l'établissement de cartes génétiques de plus en plus précises.
La technique d'irradiation va aussi être utilisée par Delbrück, qui lui s'intéresse à la nature chimique du gène. Elle permet aussi à Muller d'avancer une première estimation du nombre de gènes présents chez un organisme : il propose en 1929 que la drosophile contiendrait environ 1400 à 1800 gènes (estimation toutefois inférieure d'un ordre de grandeur au nombre obtenu à partir du séquençage complet : 13.400).
Le parcours scientifique de Muller est peu ordinaire : après avoir travaillé dans le groupe de Morgan à Columbia, puis développé sa technique de production de mutations par irradiation à l'université du Texas, il part travailler en URSS. Il y est invité par Vavilov, et doit alors affronter Lyssenko, auquel il s'oppose vigoureusement (Vavilov y laissera la vie : déporté en 1940 à l'instigation de Lyssenko, il meurt dans un camps en 1943). Il s'engage par la suite dans une brigade internationale durant la guerre civile espagnole, avant de retourner aux Etats-Unis en 1945. Il reçoit le prix Nobel de Médecine en 1946.
Muller, H.J. (1927) Artificial transmutation of the gene. Science 66, 84-87.
La première relation entre un gène et un enzyme est établie en 1902 par Archibald Garrod (St Bartholomew's Hospital, Londres), à partir d'une observation portant sur une anomalie métabolique chez l'homme : l'alcaptonurie. L'alcaptonurie est une anomalie d'excrétion, affectant le métabolisme de la tyrosine et de la phénylalanine. Les sujets atteints souffrent d'arthrite débilitante. C'est une maladie rare, dont l'incidence est estimée à 1/250000 (OMIM 203500).
Cette maladie se manifeste par le noircissement des urines lorsqu'elles sont exposées à l'air. Le noircissement est dû à la présence dans les urines d'acide homogentisique, qui est un produit intermédiaire de la dégradation de la tyrosine et de la phénylalanine. Cette substance est dégradée chez les individus normaux, mais pas chez les alcaptonuriques, chez lesquels elle s'accumule. Le sérum des premiers contient l'enzyme capable de la métaboliser (l'homogentisate 1,2 desoxygénase), mais cet enzyme n'est pas présent dans le sérum des seconds.
Garrod observe que la transmission de cette anomalie s'effectue chez l'homme en strict accord avec les lois de Mendel (selon un mode autosomal récessif), ce qui lui suggère qu'elle est due à un gène unique. Il propose donc que la déficience enzymatique soit due à une anomalie du gène responsable de la synthèse de cet enzyme. Garrod publie ces observations en 1909 dans Les erreurs innées du métabolisme, livre où il avance une explication similaire pour plusieurs maladies génétiques (albinisme, cystinurie, pentosurie). Plus généralement, il propose que chaque enzyme serait le fruit de l'activité d'un gène.
Cette observation établit pour la première fois une corrélation entre un gène et un enzyme. Cette corrélation allait être généralisée par Beadle et Tatum grâce aux mutants métaboliques de Neurospora crassa.
Garrod, A.E. (1909) Inborn errors of metabolism. Lancet 2, 1-7, 73-79, 142-148, 214-220.
Fernandez-Canon, J.M. et al (1996) The molecular basis of alcaptonuria. Nature Genet. 14, 19-24.
Le travail de Beadle est essentiellement consacré au contrôle génétique des réactions métaboliques. Les remarquables contributions de Garrod à ce domaine ne pouvaient aboutir à un programme de recherche, car des défauts génétiques décrits chez l'homme ne pouvaient évidemment pas se prêter à des expériences génétiques, envisageables uniquement sur des organismes modèles.
Beadle, en compagnie de Boris Ephrussi, choisit tout d'abord le modèle drosophile, sur lequel ils étudient la coloration des yeux (d'abord au Cal Tech, puis à Paris). Ces travaux semblent alors montrer que la synthèse des pigments responsables de cette coloration représente le résultat d'une chaîne de réactions dont chaque étape est contrôlée par un enzyme, chacun de ces enzymes étant lui-même le produit de l'activité d'un gène. Mais la complexité du système ne permet pas d'établir une conclusion définitive.
Beadle se tourne alors vers un système biochimique plus simple, décrit chez le champignon Neurospora crassa. Avec Edward Tatum, ils établissent grâce à ce micro-organisme la correspondance entre gène et enzyme (et plus généralement entre gène et polypeptide). Ces travaux sont menés à l'université de Stanford en pleine guerre mondiale, et publiés en 1941.
En 1946, Beadle prend la succession de Morgan au CalTech. De 1961 à 1968 (date de sa retraite), il est président de l'université de Chicago. Durant cette dernière période d'activité, il se consacre à l'étude du maïs.
Beadle, G.W. (1974) Recollections. Ann. Rev. Biochem. 43, 1-3.
Neurospora crassa est une moisissure facilement cultivable sur un milieu artificiel qui ne contient que du sucre, des sels minéraux, et de la biotine. Par mutagenèse artificielle (irradiation), Beadle et Tatum obtiennent des mutants incapables de se développer sur ce milieu minimal, mais qui poussent sur un milieu complémenté avec tel ou tel métabolite. En particulier, les études sur la synthèse du tryptophane, ou sur le cycle de l'ornithine, montrent que chacun de ces mutants est en réalité déficient en un des enzymes nécessaires à l'une des étapes des ces chaînes métaboliques. L'analyse génétique de ces mutants permet à Beadle et Tatum de montrer que chacune de ces déficiences segrège de façon mendélienne, et correspond donc à une mutation dans un unique gène. L'ensemble de ces observations aboutit donc à la conclusion que les gènes contrôlent la synthèse des enzymes, et que chaque protéine est codée par un gène différent. Ceci conduit Beadle et Tatum à formuler le célèbre aphorisme : "un gène <-> un enzyme", généralisé par la suite en "un gène <-> un polypeptide".
Ces travaux concrétisent alors la rencontre de la biochimie et de la génétique : la protéine vient combler la lacune qui existait entre le gène et le caractère. Beadle et Tatum s'en verront récompensés en 1958 par l'attribution du prix Nobel de physiologie et de médecine. Des travaux similaires seront par la suite produits selon cette stratégie par de nombreux autres chercheurs. Tous confirment que chaque étape des voies biochimiques est contrôlée par un gène unique, codant l'enzyme impliquée à cette étape.
Beadle, G.W. & Tatum, E.L. (1941) Genetic control of biochemical reactions in Neurospora. Proc. Natl. Acad. Sci USA 27, 499-506.
Le premier phénomène qui allait permettre de progresser dans l'identification du support de l'hérédité est celui de la transformation bactérienne, rapporté en 1928 par l'anglais Fred Griffith (1877 - 1941). Celui-ci travaille alors au laboratoire de pathologie du ministère de la santé du Royaume Uni.
Griffith décrit deux souches de pneumocoques Diplococcus pneumomiae : la souche R (rough, car lorsque cette souche est cultivée sur milieu de culture artificiel, les colonies obtenues ont un aspect rugueux) et la souche S (smooth, car les colonies ont au contraire un aspect lisse). La souche S doit son aspect à une capsule polysaccharidique qu'elle synthétise autour d'elle. Cette souche est mortelle pour la souris lorsqu'elle lui est injectée. A l'inverse, la souche R ne synthétise pas une telle capsule, et elle n'est pas nocive lorsqu'elle est injectée à une souris. On sait aujourd'hui que cette différence entre les deux souches est due à une mutation, chez la bactérie R, du gène codant l'enzyme responsable de la synthèse de la capsule.
Griffith observe tout d'abord que l'injection de bactéries S, si elles ont été préalablement tuées par la chaleur, n'est plus létale pour la souris. Pour une raison qui nous est toujours inconnue, Griffith décide alors d'injecter conjointement des bactéries S chauffées mélangées à des bactéries R vivantes. Cette fois, les souris meurent de septicémie. Les bactéries R, au contact des bactéries S tuées, ont donc acquis un caractère pathogène qu'elles ne possédaient pas précédemment. Ce phénomène a été appelé transformation bactérienne, et il a été par la suite reproduit chez plusieurs autres espèces bactériennes.
Il existe en fait plusieurs souches de pneumocoques (types I, II, ou III), discernables grâce à des tests immunologiques. Lorsque qu'une souche R de type III est injectée avec une souche S de type II inactivée, la bactérie virulente qui est ré-isolée de la souris tuée est toujours du type II. Ce changement est stable et définitif.
Ceci suggère donc qu'il existe chez les cellules un "facteur transformant", probablement libéré par la chaleur, susceptible d'être intégré par d'autres bactéries, et qui leur confère de façon héréditaire de nouvelles propriétés génétiques.
Ce phénomène représentait un test d'activité biologique, grâce auquel on pouvait envisager de déterminer la nature du matériel génétique responsable de la transformation de bactérie du type R en bactérie de type S. Griffith ne sut pas en tirer lui-même avantage, et la nature du "facteur transformant" sera élucidée plus de 10 ans plus tard par Avery et ses collègues.
Courtesy of Cold Spring Harbor Laboratory Press
 Oswald
Avery (1877 - 1955)
Oswald
Avery (1877 - 1955)La nature biochimique du matériel génétique mis en évidence par Griffith est élucidée en 1944 par les travaux de Oswald Avery, et de ses collègues (Colin McLeod, et McLyn McCarthy), qu'ils mènent à l'Institut Rockfeller de New-York. Ils reprennent les expériences de Griffith sur la transformation bactérienne, et cherchent à purifier le facteur transformant du pneumocoque.
Cette caractérisation prendra 10 ans, elle les conduit à montrer que ce facteur n'est autre que l'ADN : en effet, l'ADN extrait d'une souche S suffit à lui seul pour transformer une souche non virulente en souche virulente. La transformation des bactéries R s'effectue par incorporation de fragments d'ADN provenant des bactéries S tuées. On sait aujourd'hui que de tels fragments sont capables de rentrer dans une bactérie vivante, et de s'intégrer au chromosome de celle-ci en lieu et place de la région homologue.
L'identification de l'ADN comme principe transformant est à l'époque suffisamment extraordinaire pour nécessiter qu'une telle découverte soit étayée par des arguments indiscutables. Aussi Avery et ses collègues effectuent-ils leurs analyses avec un soin particulièrement méticuleux. Tous les contrôles alors disponibles sont testés : l'absence de protéine dans les préparations est testée par divers réactifs chimiques, leur composition chimique est analysée par des moyens chimiques ou spectrophotométrique. Enfin, l'utilisation d'enzymes montre que le pouvoir transformant réside bien dans l'ADN, puisque la DNAse anéantit ce pouvoir, alors que la RNAse ou des protéinases le laisse intact.
Avery, O.T., McLeod, C.M. & McCarthy M. (1944) Induction of transformation by a desoxyribonucleic acid fraction isolated from Pneumococcus Type III J. Exp. Med. 79, 137-158.
Malgré une accumulation croissante de preuves jusqu'au début des années 50, la communauté scientifique n'a pas accepté facilement que l'ADN puisse être le support de l'hérédité. Selon les thèses alors les plus largement acceptées, l'ADN n'est qu'une molécule simple, et donc incapable de véhiculer une information complexe. La théorie tétranucléotidique proposée par Phoebus Aaron Levene (1869 - 1940), stipulait que la structure de l'ADN est régulière et monotone, comprenant un enchaînement répétitif des 4 bases azotées. Levene était alors un des plus grands spécialistes des acides nucléiques, c'est lui qui avait identifié le désoxyribose comme un des constituants de l'ADN. Les protéines, dont on avait perçut l'immense diversité, semblaient de bien meilleurs candidats pour véhiculer une information génétique.
La découverte de Avery a été accueillie avec beaucoup de scepticisme, ils furent interprétés par beaucoup comme le résultat d'une contamination des préparations d'ADN par de faible quantité de protéines ou d'une autre substance. Et même si ces préparations d'ADN étaient absolument pures, il était possible d'envisager que cette molécule ne soit pas elle même porteuse d'information, mais qu'elle joue simplement un rôle de commutateur : toutes les informations nécessaire à la transition de la forme R vers la forme S auraient déjà été présentes dans chacune des souches, et l'ADN n'aurait alors contribué qu'à la transition d'un type vers l'autre.
Avery lui-même n'a pas véritablement cherché à imposer ses conclusions. Ainsi, l'importance fondamentale de ces travaux ne sera reconnue que tardivement, et le comité Nobel ne le retiendra pas pour l'attribution d'un prix. Lorqu'en 1953 Watson et Crick décriront la structure de l'ADN, ils ne prendront même pas la peine de citer ces travaux dans leur article.
Certains scientifiques ont cependant saisis immédiatement la portée immense des travaux d'Avery. Ce fut en particulier le cas de E. Chargaff, J. Lederberg, G. Beadle, ou de A. Lwoff.
Courtesy of Cold Spring Harbor Laboratory Press
 Erwin
Chargaff (1905 - 1992)
Erwin
Chargaff (1905 - 1992)Chargaff est un biochimiste d'origine autrichienne ayant émigré aux USA en 1934. Il fait partie des scientifiques qui saisissent immédiatement la portée immense des travaux d' Avery sur la transformation bactérienne. Lorsque Avery publie ses résultats, Chargaff dirige un laboratoire de biochimie à l'université de Columbia. Le thème central en est alors la biochimie des lipoprotéines. Quand il prend connaissance des travaux d'Avery, Chargaff comprend aussitôt que l'ADN occupe une place centrale dans les mécanismes héréditaires, et il décide de consacrer désormais les activités de son laboratoire à l'étude des acides nucléiques.
En 1950, il publie ses travaux sur le contenu en bases azotées de l'ADN chez diverses espèces, réalisés grâce aux progrès de la chromatographie sur papier. Il montre alors que le rapport A+T/C+G est variable selon les espèces, mais constant pour tous les membres d'une espèce donnée. L'ADN est donc porteur d'une certaine spécificité, cette molécule n'a pas une structure polymérique monotone, elle est donc susceptible de contenir une information. Ces travaux contribuent à répandre l'idée que l'ADN puisse être une molécule porteuse de l'information génétique.
Chargaff montre par ailleurs que le rapport C/G ou A/T est à l'inverse constant et quasiment égal à un chez toutes les espèces étudiées. Cette dernière observation sera déterminante pour l'élaboration du modèle de la structure de l'ADN par Watson et Crick quelques années plus tard.
A la suite du succès remporté par ce modèle, Chargaff est très réticent à en reconnaître l'entière paternité à ses auteurs, pour lesquels il n'a jamais eu grande estime (à la suite de sa première rencontre avec Watson et Crick, il les avait comparés à deux clowns). Il continuera à revendiquer pour son propre compte le modèle d'appariement des bases, alors qu'il n'y avait lui même jamais pensé, et n'ayant mis en évidence que les rapports égaux d'adénine et de thymine, de cytosine et de guanine.
Chargaff, E. (1950) Chemical specificity of nucleic acids and mechanism of their enzymatic degradation. Experientia 6, 201.
 Alfred
Hershey (1908 - 1997)
Alfred
Hershey (1908 - 1997)Al Hershey est, avec Delbrück et Luria, la troisième personnalité marquante du Groupe du phage. Sa première contribution importante est la mise en évidence d'une génétique des bactériophages : Il montre en effet en 1946 que peuvent apparaître des mutations chez ces organismes, caractérisés par différentes formes de plages de lyse. Ces résultats confirment donc ceux obtenus par Luria l'année précédente.
Hershey parvient aussi à détecter des phénomènes de recombinaison chez les phages : en infectant simultanément une souche bactérienne avec deux phages portant des mutations distinctes, il observe l'apparition de phages recombinants, portant soit simultanément les deux mutations, soit aucune d'entre elles. Des observations similaires seront rapportées par Delbrück.
Mais l'expérience la plus célèbre de Hershey concerne l'étude du rôle que jouent chacun des constituants du phage (ADN et protéines) dans la transmission de l'information génétique. Pour cela, il utilise - avec Martha Chase - le bactériophage T2. Leur expérience est une des premières expériences de biologie moléculaire où des isotopes radioactifs permettent de tracer des molécules. Ils utilisent un marquage isotopique différentiel de chacun des constituants du phage : du phosphore radioactif, incorporé à l'ADN, et du soufre radioactif, incorporé aux protéines de la capside. Ces phages sont utilisés pour infecter des bactéries. Immédiatement après l'infection, il est possible de séparer par agitation mécanique les bactéries des phages qui les ont infectés, et de séparer par centrifugation les particules phagiques des bactéries infectées. Lorsqu'elles sont remises en culture, ces bactéries produisent des phages. Or l'analyse de la répartition de la radioactivité montre que ces bactéries ne contiennent que du phosphore radioactif : seul l'ADN a donc pénétré dans la cellule, et cette fraction est responsable à elle seule de la reproduction du phage. C'est donc l'ADN qui détient l'information génétique.
Bien que cette expérience était dans sa conception plus grossière que celle d'Avery, son impact fut au moins aussi important. Ceci était dû en partie au fait qu'elle avait été effectuée par des membres du Groupe du phage, qui représentait alors les scientifiques les plus influents dans le domaine de la biologie moléculaire. D'autre part, ces résultats arrivaient pratiquement simultanément avec l'élucidation de la structure de l'ADN par Watson et Crick.
Hershey, A.D. (1946) Spontaneous mutations in bacterial viruses. Cold Spring Harbor Symposia on Quantitative Biology XI, 33-37.
Hershey, A.D. (1946) Mutations of bacteriophage with respect to type of plaque. Genetics 31, 620-640.
Hershey, A.D. & Chase, M. (1952) Independant functions of viral proteins and nucleic acid in growth of bacteriophage. J. Gen. Physiol. 36, 39-56.
Erwin Schrödinger fuit l'Autriche après son annexion par l'Allemagne en 1938. Sa carrière de physicien est principalement associée à la description quantique de l'atome, liée au modèle ondulatoire de la matière. Il est cependant aussi attiré par la biologie, domaine auquel il consacre en 1944 un petit livre : Qu'est ce que la vie ? Il y développe des idées d'une extraordinaire clairvoyance :
- il décrit le matériel génétique comme un "cristal apériodique". On sait aujourd'hui que l'ADN présente effectivement globalement deux ordres de symétrie (rotationnel et translationnel), cette régularité l'apparente bien à un cristal. Mais dans le détail, l'enchaînement des paires de bases n'est pas exactement répétitif, ce cristal est apériodique.
- il insiste sur la notion de programme, de code génétique : "ces chromosomes (...) qui contiennent sous la forme d'une espèce de code le modèle intégral du développement futur de l'individu et de son fonctionnement dans l'état adulte".
- enfin, il expose sa profonde conviction que le problème du gène est nécessairement accessible à l'expérimentation. En particulier, son appréciation des travaux de Delbrück sur la mutagenèse induite par les rayons X est remarquable.
De nombreux biologistes fameux, parmi lesquels Watson, Crick, ou Wilkins ont reconnu que la lecture de Qu'est ce que la vie ? a été décisive pour leur engagement dans la recherche en biologie.
Schrödinger, E. (1944) Qu'est ce que la vie ? (trad. française : Points/Seuil, 1986)
 Max
Delbrück (1906 - 1981)
Max
Delbrück (1906 - 1981)L'influence de Delbrück, bien que largement reconnue, est difficile à expliquer, car son œuvre n'est pas associée à une avancée scientifique majeure. C'est essentiellement sa rigueur et son charisme scientifiques qui en ont fait le leader de ce qui allait devenir la biologie moléculaire.
Max Delbrück est né à Berlin, et suit tout d'abord un cursus de physicien. Après sa thèse, obtenue en 1930, il rejoint le laboratoire de Niels Bohr à Copenhague. L'influence de ce dernier le pousse alors à s'intéresser à la biologie, et plus précisément à la nature du gène. De retour à Berlin en 1932, Delbrück réalise ses premiers travaux sur les propriétés physiques du gène.
La première expérience menée par Delbrück est révélatrice de sa formation : de même que les physiciens étudient l'atome de façon indirecte grâce à un bombardement particulaire, Delbrück décide d'étudier le gène par le biais de l'effet que des rayonnements induisent sur celui-ci. Comme l'avait montré Muller, la fréquence de mutations peut être accrue par un rayonnement du type rayons X ou gamma. Delbrück cherche alors à préciser les propriétés du gène en étudiant les variations du taux de mutation en fonction de l'énergie du rayonnement utilisé. L'analyse de la fréquence des mutations obtenues permet à Delbrück en 1935 d'estimer la dimension d'un gène à un volume d'environ 10 distances atomiques de côté, ne contenant donc qu'un millier d'atomes environ. Delbrück interprète les mutations en termes quantiques : il propose que chaque mutation représente un saut quantique entre deux états stables du gène. Bien que par plusieurs aspects ces conclusions soient erronées, le mérite de Delbrück est d'avoir montré que le vivant pouvait être approché par les outils et les démarches du domaine de la physique. Schrödinger allait accorder une grande importance à ces travaux dans son livre Qu'est ce que la vie?
A partir de 1937, Delbrück travaille aux Etats-Unis, au California Institute of Technology (Pasadena). Convaincu que le meilleur modèle d'étude du vivant doit être le plus simple possible, il choisit le bactériophage, dont la très faible taille l'assimile à un "gène pur". Ce matériel devrait par conséquent se prêter plus facilement à l'étude des mécanismes de réplication. Les premiers travaux de Delbrück sur le phage ont porté sur l'analyse de sa cinétique de croissance. Avec Emory Ellis, ils montrent que celle-ci s'effectue de façon discontinue, des phases de brusque multiplication alternant avec des phases stationnaires. La phase de multiplication correspond à la phase de lyse cellulaire libérant les phages qui viennent d'être produits au cours du cycle d'infection précédent. Cette description de la "croissance en une étape" eut un retentissement significatif : la façon très rigoureuse de mener les expériences, leur analyse statistique soignée, le détachement total par rapport à toute idée préconçue, étaient remarquables, et ces caractéristiques allaient marquer tous ses travaux ultérieurs. En 1940, Delbrück s'installe à l'université Vanderbilt à Nashville, puis retourne au Cal Tech à partir de 1947.
Delbrück va devenir le chef de file de ce qui allait devenir le Groupe du Phage, qui regroupait l'ensemble des chercheurs ayant choisi cet organisme comme modèle d'étude. Ce groupe informel allait s'étoffer avec en particulier Salvador Luria (à partir de 1941) et Alfred Hershey (à partir de 1943). Des cours seront organisés chaque année (pendant 26 ans) sur cet organisme au Cold Spring Harbor Laboratory.
La mise au point du test de fluctuation par Delbrück et Luria en 1943 font entrer la bactérie dans le monde de la génétique moderne. En 1946, Delbrück toujours, et indépendamment Hershey, rapportent les premiers indices témoignant de recombinaisons possibles entre phages portant des mutations distinctes. Cela allait aboutir aux premières cartes génétiques de phages.
Par la qualité de ses travaux, Delbrück et le Groupe du phage allaient acquérir une réputation considérable. De son vivant même, un livre sera consacré à Delbrück à l'occasion de son 60ème anniversaire. Enfin, Luria, Hershey, et Delbrück recevront en 1969 le prix Nobel de médecine pour l'ensemble de leurs travaux.
Après 1953, et la solution de la structure de l'ADN qui apportait enfin une réponse au problème de la réplication du gène, Delbrück se détourne du phage pour se consacrer à l'étude de la réponse à la lumière. Il choisit pour cela ce qu'il pense être l'équivalent du phage pour ce problème : le champignon Phycomyces , sur lequel il travaillera encore 25 ans (il y aura aussi un Groupe du Phycomyces).
Ellis, E. & Delbrück, M. (1939) The growth of bacteriophage. J. Gen. Physiol. 22, 365-384.
Luria, S.E. & Delbrück, M. (1943) Mutations from virus sensitivity to virus resistance. Genetics 28, 491-511.
Schrödinger, E. (1944) Qu'est ce que la vie ? (trad française : Points/Seuil, 1986)
Cairns, J., Stent, G.S. & Watson, J.D. (1966) Phage and the origins of molecular biology. CSHL Press.
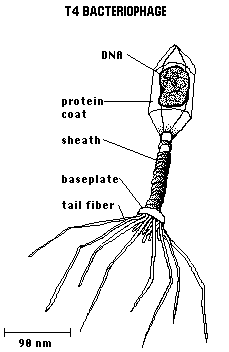 Le
bactériophage
Le
bactériophageLes bactériophages sont mis en évidence indépendamment en 1915 par l'anglais Frederick Twort (Londres) et par un franco-canadien, Felix d'Hérelle (Institut Pasteur). Ils sont alors caractérisés comme des entités de taille extrêmement faible (capables de traverser des filtres très fins qui par ailleurs retenaient les bactéries), susceptibles de détruire des bactéries.
Les bactériophages sont des entités très simples, constituées d'une tête et d'une queue qui forment une capside protéique. Dans la tête est logée une molécule d'ADN. Les bactériophages ne sont pas à proprement parler des êtres vivants, car ils sont incapables de se reproduire de façon autonome, et ils exigent pour cela d'interagir avec une bactérie qu'ils infectent. Cette infection s'effectue par fixation du phage sur une bactérie au niveau de récepteurs spécifiques, injection de leur matériel génétique, et détournement de la machinerie enzymatique de la bactérie hôte pour la production de plusieurs centaines de nouveaux phages.
A l'époque ou ce système phage/bactérie commence à percer, il représente un système de reproduction fascinant de par son apparente simplicité, sa facilité et sa rapidité de mise en œuvre (un cycle lytique s'effectue en 20 minutes environ). Il semble alors constituer le modèle idéal pour s'attaquer au problème du gène et de sa réplication. De plus, avec ces organismes, ce sont des millions de "descendants" qui peuvent être obtenus à chaque "génération" (alors que le modèle favori jusque là des généticiens, la drosophile, ne permet d'examiner que quelques centaines de descendants par génération). Le nombre de mutations détectables s'en trouve accru d'autant, ce qui permettra en particulier d'observer et d'étudier des recombinaisons intragéniques. Ce sont ces organismes extrêmement simples, qui vont apporter les preuves ultimes que l'ADN est le support de l'hérédité (expérience de Hershey et Chase).
En 1944, le nombre de chercheurs impliqués dans la recherche sur les bactériophages s'est accru dans de telles proportions que Delbrück est amené à proposer un "Traité des Phages", qui encourage les chercheurs de cette communauté à utiliser les mêmes systèmes bactériophages/bactéries, afin que l'ensemble des résultats obtenus soient comparables et intégrables. Les systèmes alors choisis sont les phages de la série T (T1 à T7), et leur bactérie hôte, E. coli.
 Salvador
Luria (1912 - 1991)
Salvador
Luria (1912 - 1991)Salvador Luria est né à Turin. Médecin de formation, il se tourne néanmoins vers la physique, et travaille tout d'abord dans le laboratoire d'Enrico Fermi. Puis il se tourne vers la biologie, et commence en particulier à s'intéresser au bactériophage. La montée de l'antisémitisme le conduit à quitter l'Italie pour la France en 1938, d'où il fuit l'invasion allemande en 1940 pour se réfugier aux Etats-Unis. Il travaille tout d'abord à l'Université Columbia à New-York, avant d'obtenir un poste à l'université de l'Indiana à Bloomington.
A partir de 1941, il collabore avec Delbrück sur le système bactérie/phage. Leur première contribution majeure est le développement du test de fluctuation. Ce teste avait pour but d'identifier l'origine de bactéries résistantes aux bactériophages, qui peuvent apparaître lorsque ces organismes sont mis en présence. Luria et Delbrück furent les premiers à poser clairement ce problème, et à y répondre par des arguments expérimentaux (1943). Deux hypothèses étaient susceptibles d'expliquer l'apparition de cette résistance : soit le phage induit lui même la résistance (vision lamarckienne), soit les bactéries mutantes pré-existent à la confrontation avec le phage, et sont sélectionnées par la pression sélective que celui-ci exerce (vision darwinienne). Dans le premier cas, le nombre de bactéries résistantes devrait être uniforme à chaque expérience, le nombre moyen de bactéries résistantes représentant simplement le produit du nombre de bactéries initiales par la probabilité qu'une bactérie développe la résistance au virus avant que celui la tue. Dans le second, il devrait être aléatoire, dépendant uniquement du nombre de bactéries mutantes déjà présentes dans la population étudiée avant l'introduction du phage. Lorsque Luria testa ces deux hypothèses, il observa un nombre de bactéries résistantes très variable selon les expériences, ce qui validait la seconde hypothèse. Ce résultat constituait la première preuve que les bactéries pouvaient subir des mutations, il consacrait la naissance de la génétique bactérienne.
Avec Anderson, Luria obtient en 1942 les premières photographies en microscopie électronique de bactériophage. Enfin, Luria montre en 1946 que peuvent apparaître des mutations chez les phages : des phages incapables d'infecter une souche bactérienne résistante peuvent par exemple acquérir par mutation la capacité d'infecter cette souche.
Dans les années suivantes, Luria poursuit ses travaux au sein du Groupe du phage. Ceci le conduira à partager le prix Nobel de physiologie et médecine en 1969 avec Delbrück et Hershey. Aujourd'hui, sa mémoire est toujours présente dans les laboratoire, puisque le milieu L a conservé l'initiale de son nom.
Luria, S.E. & Delbrück, M. (1943) Mutations of bacteria from virus sensitivity to virus resistance. Genetics 28, 491-511.
Luria, S.E. & Anderson, T.F. (1945) The identification and characterization of bacteriophages with the electron microscope. Proc. Natl. Acad. Sci. USA 28, 127-130.
Luria, S.E. (1945) Mutations of bacterial viruses affecting their host range. Genetics 30, 84-99.
La structure en double hélice de l'ADN est
élucidée par Watson et Crick en 1953. Watson a décrit dans son livre passionnant
La double hélice (1968) le récit de la formidable
découverte réalisée avec Crick . Les deux
chercheurs disposent alors des éléments suivants : (i)
la composition chimique de l'ADN (désoxyribose, bases
azotées, et groupements phosphate) ; (ii) les clichés de
diffraction aux rayons X d'ADN cristallisé, clichés dus
principalement à Rosalind Franklin et
Maurice Wilkins du King's College. Ces
clichés montrent une figure en croix, caractéristique
des structures en hélice ; (iii) les travaux de Erwin Chargaff,
qui avaient montré que pour toute molécule d'ADN, le
nombre de molécules d'adénine est égal au nombre
de molécules de thymine, et que celui de cytosine est
égal à celui de guanine ; (iv) les analyses en
microscopie électronique, qui avaient montré que le
diamètre de la molécule d'ADN est de 20 Å, ce qui
suggérait que cette molécule comportait deux
chaînes de désoxyribose-phosphate.
C'est en élaborant successivement plusieurs modèles moléculaires que Watson et Crick réussissent à proposer une structure qui satisfasse à l'ensemble des données cristallographiques et biochimiques alors disponibles. Cette structure est aujourd'hui connue de tous, elle est devenue l'emblème de la biologie moléculaire : deux brins constitués des groupements phosphates et des sucres forment une double hélice où les orientations de chacun des brins sont opposées. Sur les sucres de chacun des deux brins sont liées les bases azotées, chaque base d'un brin étant maintenue en vis-à-vis d'une base de l'autre brin par des liaisons hydrogène. Une cytosine fait toujours face à une guanine, et une adénine à une thymine. Les deux brins d'une molécule d'ADN sont dits complémentaires.
Crick, Watson, et Wilkins reçurent en 1962 le prix Nobel pour ces travaux, qui a été qualifiée par Peter Medawar de "la plus grande réussite scientifique de notre siècle". Rosalind Franklin aurait vraisemblablement été associée à ce prix si la maladie ne l'avait prématurément emportée.
L'ordre des bases le long de la molécule définit l'information génétique portée par l'ADN. Les possibilités offertes par l'agencement des quatre bases de l'ADN sont immenses (4n pour une séquence de n bases, soit par exemple plus d'un million de combinaisons pour une séquence de 10 bases).
L'ordre des bases nucléotidiques détermine l'enchaînement des acides aminés des protéines (par l'intermédiaire des processus de transcription et de traduction). Une structure à une dimension détermine donc des structures à 3 dimensions. Ce paradoxal "gain d'information" est réalisé spontanément, par le reploiement des protéines au fur et à mesure de leur synthèse, en fonction des contraintes thermodynamiques imposées par leur séquence et par le milieu dans lequel elles sont synthétisées.
Les êtres vivants disposent donc d'un système informatif à une dimension, entièrement compris dans une séquence linéaire, qui peut être facilement copié ou dupliqué. De cette séquence linéaire découle celle des protéines, qui adoptent spontanément la structure à trois dimensions nécessaire à leur fonction.
La propriété auto-réplicative des gènes constituait un phénomène mystérieux, qui avait suscité de nombreuses théories explicatives. Celle qui s'approchait le plus de la réalité fut sans doute proposée par Pauling et Delbrück en 1940 : probablement, une telle opération pouvait s'effectuer par l'intermédiaire d'une structure intermédiaire, un moule en négatif, dont le moulage produira à son tour une structure identique à l'original.
Dans la publication de leur modèle, Watson et Crick mentionnent : "il n'a pas échappé à notre attention que l'appariement spécifique des bases que nous avons proposé suggère immédiatement un mécanisme possible de réplication pour le matériel génétique". Cet appariement suggère en effet le mécanisme par lequel chacun des brins complémentaires de l'ADN peut être à la fois matrice et participant pour la constitution de deux nouvelles molécules d'ADN : après séparation des deux brins, chacun d'eux est utilisé comme matrice pour la synthèse du brin complémentaire, ce qui aboutit à la formation de deux molécules identiques entre elles et identiques à la molécule initiale.
 James
Dewey Watson (1928 - )
James
Dewey Watson (1928 - ) Jim Watson est né à Chicago le 6 avril 1928. Très précoce, il entre à l'université de Chicago à l'âge de 15 ans. Il prépare sa thèse entre 1948 et 1950 dans le laboratoire de Salvador Luria, dont il est le premier étudiant. Il est donc bien informé des travaux menés par le groupe du phage. Il effectue une courte période post-doctorale au Danemark, avant de rejoindre en 1951 Francis Crick à Cambridge au laboratoire Cavendish (dirigé par Max Perutz et Lawrence Bragg). En 1953, Crick et Watson élucident la structure en double hélice de l'ADN . A partir de 1953, Watson est Senior Research Fellow au Californian Institute of Technology , jusqu'en 1956, où il devient professeur à l'université de Harvard.
En 1962, le prix Nobel de physiologie et de médecine est attribué à James Watson, Francis Crick et Maurice Wilkins, pour "leur découverte concernant la structure moléculaire des acides nucléiques et son importance concernant le transfert d'information dans les systèmes vivants". La même année est attribué le prix Nobel de chimie à Max Perutz et John Cowdery Kendrew pour "leurs études de la structure des protéines globulaires". Ces deux prix couronnent l'application de la diffraction des rayons X à la détermination de structures de molécules biologiques.
En 1968, Watson prend la direction du Cold Spring Harbor Laboratory , où il développe en particulier la recherche sur les bases moléculaires du cancer. Il dirige alors parallèlement deux laboratoires (l'un à Harvard, l'autre à CSH), jusqu'en 1977 où il renoncera à celui d'Harvard. En 1988, dans le cadre du Projet Génome Humain , lui est confiée la direction du programme Génome Humain du National Institutes of Health (qui allait devenir l'année suivante le NCHGR : National Center for Human Genome Research of the National Institutes of Health). Il en démissionne en 1992, lorsque le NIH manifeste la volonté de prendre des brevets sur des séquences partielles de cDNA humains. Il est actuellement président du CSHL.
Watson, J.D. & Crick, F.H.C. (1953) A structure for desoxyribose nucleic acid. Nature 171, 737-738.
Watson, J.D. & Crick, F.H.C. (1953) Genetical implications of the structure of deoxyribonucleic acid. Nature 171, 964-967.
Watson J.D. (1968) La double hélice, Hachette/Pluriel.
Recombinant DNA, (1992) Watson, J.D., Gilman, M., Witkowski, J. & Zoller, M. Scientific American Books.
Molecular biology of the cell, (1994) Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. & Watson, J.D. Ed. Garland Publishing.
 Francis
Harry Compton Crick (1916 - 2004)
Francis
Harry Compton Crick (1916 - 2004)Francis Crick est physicien de formation. Il travaille tout d'abord à l'University College de Londres, sur la viscosité de l'eau à haute pression et haute température. Pendant la guerre, il est embauché par l'Amirauté pour la mise au point et la détection de mines magnétiques. Une fois la guerre finie, il s'oriente vers la cristallographie, au sein du laboratoire Cavendish, dirigé par Max Perutz et Lawrence Bragg. C'est là que le rejoint Jim Watson en 1951. Crick a alors 35 ans, et il prépare toujours sa thèse. Watson et Crick se consacrent alors à la structure de l'ADN, pour laquelle ils proposent un modèle en 1953. Ces travaux seront couronnés par un prix Nobel attribué en 1962.
Par la suite, toujours au laboratoire Cavendish, Crick s'attache avec Sidney Brenner au déchiffrage du code génétique. Après que ce code est élucidé en 1966, Crick se tourne vers la biologie du développement. Enfin, en 1976, Crick rejoint le Salk Institute for Biological Studies, près de San Diego, où il se consacre à la neurobiologie.
Crick a apporté d'importantes contributions à plusieurs domaines de la biologie moléculaire : c'est à lui que l'on doit le dogme central de la biologie, qui stipule que le flux d'information depuis les acides nucléiques vers les protéines est à sens unique. Par ailleurs, il est un des premiers auteurs à proposer en 1968 que le matériel génétique des premiers organismes vivants ait été de l'ARN. Enfin, en 1980, il contribue à l'élaboration de la notion d'ADN égoïste.
Watson, J.D. & Crick, F.H.C. (1953) A structure for desoxyribose nucleic acid. Nature 171, 737-738.
Watson, J.D. & Crick, F.H.C. (1953) Genetical implications of the structure of deoxyribonucleic acid. Nature 171, 964-967.
Crick, F.H.C. (1968) The origin of the genetic code. J. Mol. Biol. 38, 367-379.
Crick, F.H.C. (1970) Central dogma of molecular biology. Nature 227, 561-563
Orgel, L.E. & Crick, F.H.C. (1980) The selfish DNA : the ultimate parasite. Nature 284, 604-607.
Courtesy of Cold Spring Harbor Laboratory Press
 Rosalind Franklin (1920 - 1958)
Rosalind Franklin (1920 - 1958)Née en 1920, Franklin suit ses études à l'université de Cambridge. En 1947, elle travaille dans un laboratoire parisien, jusqu'en 1951, où elle rejoint le King's College, pour travailler sur la structure de l'ADN. Ses rapports avec ses collègues sont médiocres, et Watson en a brossé un portrait caricatural dans La double hélice. C'est toutefois à elle que l'on doit tous les clichés de diffraction aux rayons X de l'ADN, qui permettent à Watson et Crick de proposer leur modèle en 1953. Elle reconnaît d'autre part deux formes distinctes pour les cristaux d'ADN, qu'elle nomme forme A (forme compactée) et forme B (forme hydratée). Ces travaux sont publiés dans le même numéro de Nature que celui décrivant le modèle de Watson et Crick.
En mars 1953, Franklin quitte le King's College pour le Birbeck College. Elle se consacrera désormais à l'étude du virus de la mosaïque du tabac, dans le laboratoire du professeur Bernal. Une leucémie l'emportera à l'âge de 38 ans.
Franklin, R.E. & Gosling R.G. (1953) Molecular configuration in sodium thymonucleate. Nature 171, 742.
Franklin, R.E. & Gosling R.G. (1953) Evidence for 2-chain helix in crystalline structure of sodium desoxyribonucleate. Nature 172 156.
Klug, A. (1968) Rosalind Franklin and the discovery of the structure of DNA. Nature 219, 808-844.
 Linus
Pauling (1901 - 1994)
Linus
Pauling (1901 - 1994)Linus Pauling (1901 - 1994) fut sans doute le plus grand chimiste de ce siècle, qui aborda avec succès des domaines variés tels que la physique quantique, la structure des cristaux, ou la biologie moléculaire. Ses premières contributions essentielles concernent la liaison chimique, et il est en particulier le premier à reconnaître l'importance des liaisons faibles. En 1940, avec Delbrück, il développe l'idée que la complémentarité stéréospécifique est à la base des interactions moléculaires en biologie. Il publie deux livres qui ont révolutionné la chimie : La nature de la liaison chimique (1939), et Chimie générale (1947). Plus que tout autre, il contribue à faire de la structure des molécules le thème essentiel de la biochimie. Il élucide la structure de plusieurs centaines de substances minérales, et il est le découvreur de certaines structures protéiques, telles que l'hélice alpha et le feuillet beta;. Il aussi le premier à apporter une explication moléculaire à une maladie génétique : l'anémie falciforme, pour laquelle il propose comme origine une altération de l'hémoglobine.
L'ensemble de ces travaux lui vaut un premier prix Nobel en 1954. Un prix Nobel de la paix lui est aussi attribué en 1962, en raison de ses engagements pacifistes en faveur du désarmement (seuls trois autres chercheurs ont reçu deux prix Nobel : Marie Curie, Antoine-Henri Becquerel, et Frederick Sanger). Là encore, il fait figure de pionnier, puisqu'il est le premier à avancer que les radiations puissent être dangereuses pour le patrimoine génétique.
Pauling est aussi à l'origine de l'utilisation des séquences de protéines pour la reconstruction d'arbres phylogéniques, et il propose avec Emile Zuckerkandl, en 1965, le concept d'horloge moléculaire. Il travaille aussi sur la structure de l'ADN, et a été à ce titre le concurrent le plus sérieux de Watson et Crick . En 1953, il publie ce qu'il pense en être la structure : trois chaînes de riboses phosphates s'enroulant étroitement l'une autour de l'autre, et les bases azotées pointant radialement vers l'extérieur de ce faisceau. Cette idée totalement erronée s'explique en partie par le fait que Pauling n'avait pas accès aux résultats récents de Rosalind Franklin .
A partir des années 70, il s'intéresse aux antioxydants, en particulier la vitamine E et la vitamine C. Il accorde à ces molécules d'importantes vertus anti-cancérigènes et anti-vieillissantes. Ceci le conduit vers un singulier engouement pour la vitamine C, dont il prône la consommation sans modération...
Pauling, L. (1948) Nature of forces between molecules of biological interest. Nature 161, 707-709.
Pauling, L. & Delbrück, M. (1940) The nature of the intermolecular forces operative in biological processes. Science 92, 77-79.
Pauling, L., Itano, H.A. & Singer, S.J. (1949) Sickle cell anemia, a molecular disease. Science 110, 64-66.
Pauling, L., Corey, R.B. & Branson, H.R. (1951) The structure of proteins : two hydrogen-bonded helical configurations of the polypeptide chain. Proc. Natl. Acad. Sci. USA 37, 205-211.
Pauling, L. & Corey, R.B. (1953) A proposed structure for the nucleic acids. Proc. Natl. Acad. Sci. USA 39, 84-97.
Zuckerkandl, E. & Pauling, L. (1965) Molecules as documents of evolutionary history. J. Theoret. Biol. 8, 357-366.
Cairns, J., Stent, G.S. & Watson, J.D. (1992) Phage and the origins of molecular biology, Cold Spring Harbor Laboratory Press
Crick, F. (1988) Une vie à découvrir, Odile Jacob
Gros, F. (1986) Les secrets du gène, Points/Seuil
Jacob, F. (1981) Le jeu des possibles, Le Livre de Poche
Jacob, F. (1970) La logique du vivant, Gallimard
Judson, H.F. (1996) The eighth day of creation, Cold Spring Harbor Laboratory Press
Mayr, E. (1989) Histoire de la biologie, Fayard
Morange, M. (1994) Histoire de la biologie moléculaire, La Découverte
Schrödinger E. (1986) Qu'est ce que la vie ? Points/Seuil
Watson J.D. (1984) La double hélice, Hachette/Pluriel
© généthon
{kind=link}